Heterotrimeric GTP-binding proteins (G proteins) are key mediators in a majority of signaling systems that relay hormonal and sensory stimulation of specific cell surface receptors to intracellular effectors. Our reserch focuses on the visual transduction cascade in vertebrate rod photoreceptors that has served for many years as a paradigm for G protein signaling.
In rod photoreceptors, illuminated rhodopsin (R*) stimulates GTP-GDP exchange on the α-subunit of the retinal G protein, transducin (Gt1). Activated Gαt1 molecules, Gαt1GTP, dissociate from the Gβ1γ1-subunits and R*, and stimulate the effector enzyme, cGMP phosphodiesterase (PDE6), by displacing the inhibitory γ-subunits (Pγ) from the PDE catalytic core (PDE6αβ). cGMP hydrolysis by active PDE6 results in closure of cGMP-gated channels in the plasma membrane . The turn-off phase of the visual signal is determined by reactions controlling the lifetimes of photoexcited rhodopsin (R*) and activated transducin. The ability of R* to continue to activate transducin is blocked by phosphorylation of the C-terminal Ser and Thr residues by rhodopsin-kinase and binding of arrestin to phosphorylated R*. The lifetime of Gαt1GTP depends upon its GTPase activity. Hydrolysis of GTP switches the Gαt1 molecule to the inactive GDP-bound conformation and allows re-inhibition of PDE6αβ by Pγ. Currently, we are concentrating our efforts on two major projects.
I. Regulation of photoreceptor cGMP phosphodiesterases (PDE6).
Phosphodiesterases of cyclic nucleotides are critical enzymes modulating cellular levels of cAMP and cGMP. Eleven families of PDEs have been identified in mammalian tissues based on primary sequence, substrate selectivity, and regulation. Photoreceptor rod and cone PDEs comprise the PDE6 family. In various species, rod PDE6 is composed of two homologous catalytic α- (PDE6A) and β-subunits (PDE6B)) and two copies of an inhibitory γ subunit (Pγ). Cone PDE6 is composed of two identical catalytic PDE6C subunits each bound to a cone-specific inhibitory Pγ subunit. PDE6 catalytic subunits contain three structural domains: N-terminal regulatory GAFa and GAFb domains and the conserved C-terminal catalytic domain.
The failure of functional expression of PDE6 in various systems has impeded the understanding of the structural basis of PDE6 function. For a number of years, we have analyzed PDE6 function through characterization of chimeric enzymes between PDE6 and the related PDE5 enzyme, which is readily attainable as recombinant protein.
Our mutational analysis of PDE6 residues using PDE5/PDE6 chimeric templates has revealed the key determinants of dimerization and exceptional catalytic efficiency of PDE6, and identified essential Pγ contact residues.
Recently we have generated a fully functional chimeric PDE5/PDE6 catalytic domain (PDE5/6cd) that was potently inhibited by Pγ. In collaboration with our colleagues from the Biochemistry Department, L. Gakhar and S. Ramaswamy, we solved the structures of PDE5/6cd in complex with sildenafil (Viagra) or in complex with 3-isobutyl-1-methylxanthine and the Pγ-inhibitory peptide, Pγ70-87. (Barren et al., 2009) These structures yielded important molecular details on the interaction of the Pγ C-terminus with the PDE6 catalytic subunits and the relationships between the Pγ- and drug-binding sites at the catalytic pocket. The structures show that the C-terminus of Pγ occludes the opening of the catalytic pocket and blocks access of cGMP to the active site.
The movie below details the Pγ C-terminus blocking the catalytic site of PDE6
The structure of the PDE-bound Pγ C-terminus also suggests Pγ conformational change upon activation by transducin. Based oncomparison of the Pγ C-terminus conformations in the PDE5/6cd complex and the transition-state complex of Gat (Slep et al, 2001), we proposed that GatGTP binding induces a conformational change encompassing Pγ residues 71-77 and results in a hinge-like rigid-body movement of Pγ-78-87 away from the PDE6 catalytic pocket leading to PDE6 activation.
The movie below demonstrates the proposed Pγ conformational change on PDE6 activation by transducin
In a search for a system for expression and analysis of PDE6, we recently explored the utility of transgenic frogs Xenopus laevis. We ectopically expressed the EGFP-fusion protein of human cone PDE6C in rods of Xenopus laevis and investigated subcellular localization, membrane diffusion, and biochemical properties of the recombinant PDE6 enzyme (Muradov et al. 2009).
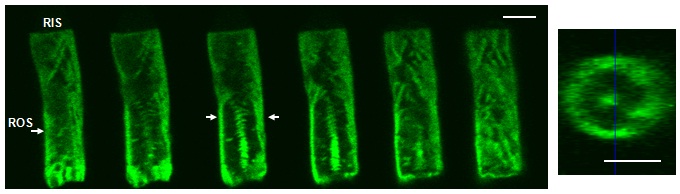
The movie below shows projections of EGFP PDE6-C retina.
The pattern of EGFP-PDE6C fluorescence in transgenic Xenopus rods indicates that the enzyme is concentrated at the disc rim region and incisures (for details see Muradov et al., 2009).
Transgenic Xenopus laevis appear to represent a unique expression system for PDE6 well-suited to address such outstanding questions as (1) properties of the individual rod PDE6A and PDE6B catalytic subunits and the functional significance of their heterodimerization (see for example Muradov et al., 2010); (2) PDE6 assembly and traffic in photoreceptor cells; (3) analysis of differences between rod and cone PDE and their potential contribution to the distinct physiology of rods and cones, and (4) mechanisms of visual diseases caused by PDE6 mutations.
II. Molecular Mechanisms of photoreceptor G-protein signaling.
The overall goal of this research program is to gain insights into the molecular mechanisms of the GTP-binding protein, transducin, in photoreceptor cells.
Our current focus is on two key questions. What mechanisms govern bi-directional translocation of transducin between the inner and outer segments in rods? What are the signaling properties of cone transducin, and how do they contribute to the distinct physiology of cone photoreceptors?
Three major visual signaling proteins, transducin, arrestin, and recoverin undergo bi-directional translocations between the outer segment and inner compartments of rod photoreceptors in a light-dependent manner.The light-dependent translocation of proteins is believed to contribute to adaptation and neuroprotection of photoreceptor cells. However, the physiological significance and mechanisms of light-controlled transducin translocation are not fully understood. Studies from several laboratories, including our lab, have supported a simple diffusion model of transducin translocation in response to light. According to the model, the activation of transducin by photoexcited rhodopsin causes dissociation of Gt subunits allowing them to diffuse into the inner segment as long as the Gt activation rate exceeds the capacity of the rod RGS9 GAP complex to inactivate Gαt1GTP. The mechanisms of transducin transport from the rod inner segment to the outer segment are largely unknown.
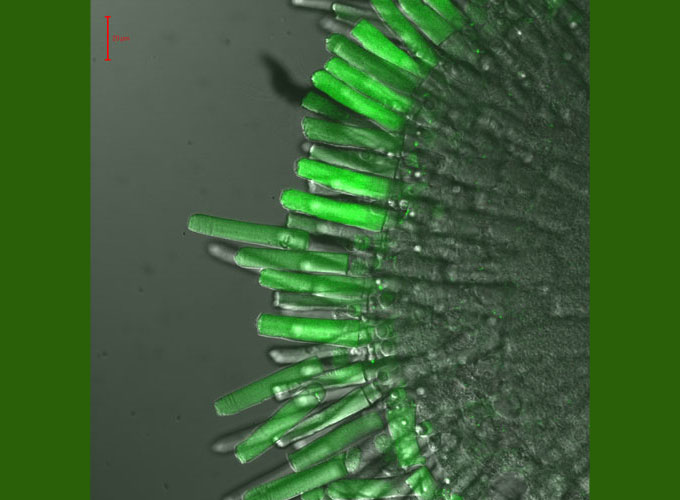
As one of the approaches to study tranducin transport and targeting, we are utilizing transgenic Xenopus laevis expressing EGFP-fused Gαt1 and mutant forms in rod photoreceptors. Transgenic Xenopus models have been designed to probe the roles of transducin/rhodopsin interactions and lipid modifications in transducin targeting, membrane mobility, and inter-disc transfer. Light-dependent compartmentalization, translocation, and mobilities of transducin and mutants are examined with EGFP imaging, immunofluorescence, and Fluorescence Recovery After Photobleaching (FRAP) analysis of lateral and longitudinal diffusion (for details see our recent study Kerov and Artemyev, 2010).
A piece of retina of a transgenic tadpole expressing EGFP- Gαt1. EGFP-fluorescence/DIC overlay.